
Authors:
I.Torshin
O.Gromova
O.Limanova
Lithium Ascorbate, a component of Normotim. It’s the safest lithium salt, which is 12 times safer than lithium carbonate. Lithium Ascorbate is also 10 times more effective than lithium carbonate in low doses.
It is well known that the complex of nerve and hormonal signals under stress stimulates the secretion of adrenaline, noradrenaline, and cortisol by the adrenal glands. Much less attention has been given to the roles of intermediate signaling proteins that mediate the effects of “stress hormones,” neurotransmitters, and other signaling molecules. This paper presents the results of systematization of studies of the glycogen synthase kinases GSK-3α and GSK-3β, excessive activity of which aggravates the course of chronic stress and has a negative impact on the survival of neurons and adaptation processes. Lithium ions are natural inhibitors of the excess activity of both GSK-3, which is partially responsible for the normothymic and antidepressant effects of lithium-based preparations. Administration of lithium salts based on organic anions is the safest and most effective way to replenish lithium deficiency in the body. We considered the prospects of using lithium ascorbate to increase the adaptation reserves in the body. Lithium Ascorbate is one of the main components of Normotim from Normopharm, which is now actively used in the treatment of stress and depression.
Stress is something that accompanies a person throughout their entire life and is a kind of “test” for the body’s adaptation to environmental factors. When the body’s resistance to stress decreases, the so-called “exhaustion stage” occurs, which, when chronic, promotes the development of various diseases. According to Hans Sellier, the founder of stress physiology theory, “it is not the stress itself that kills us, but our reaction to it”. Therefore, there is a distinction between “eustress” (the term implies the correct reaction of the body to stress) and “distress” (pathological reaction to stress, leading to exhaustion, depression, excessive aggression, as well as damage to various systems of the body). The word “stress” is commonly used to refer to distress.
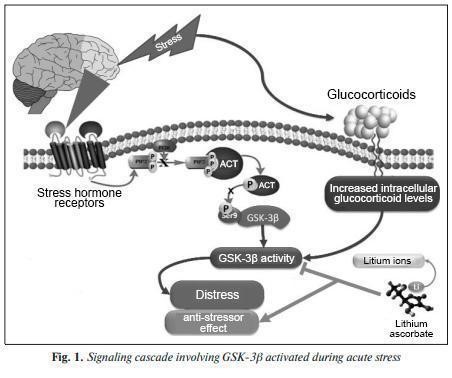
Chronic increases in the levels of mediators of the neuroendocrine response to stress (adrenocorticotropic hormone, cortisol, noradrenaline, dopamine, etc.) have a detrimental effect on the brain by disrupting metabolism, neuroplasticity, and neuron survival. In turn, loss of neuroplasticity negatively affects perception of events and stress adaptation. The brain structures involved in response to chronic distress (cortex, hippocampus, hypothalamus, and amygdala) are subjected to its maladaptive influence with subsequent disruption of the hypothalamic-pituitary-adrenal axis, adversely affecting the hormonal, immune, reproductive, hematopoietic, and antioxidant systems of the body. Even moderate chronic stress leads to the appearance of such symptoms of neurodegeneration as impaired synaptic transmission, accumulation of β-amyloid and hyperphosphorylation of τ-protein. These effects appear on the basis of excessive activation of glutamate NMDA receptors, which leads to excitotoxic death of neurons. For this reason, the problem of drugs aimed at maintaining stress resistance in the body has been opened up in neuropharmacology. One of the factors contributing to the development of distress is the deficiency of micronutrients, and lithium in particular. The results of experimental studies indicate adaptogenic, normothymic, neurotrophic, and neuroprotective effects of lithium. In very low doses (fractions of a milligram), lithium promotes acceleration of neuronal growth and increases their resistance to oxidative stress and long-term potentiation in the hippocampus. Lithium administration in patients with bipolar disorder has been shown to stimulate an increase in the volume of gray matter in the brain. Unlike synthetic tranquilizers and sedatives that directly affect certain neuroreceptors, lithium salts restore neuropsychic activity through multimodal action, which is a more physiological way of counteracting stress. Excessive activation of adrenal and cortisol-dependent systems is regulated by lithium through inhibition of glycogen synthase kinase-3β (GSK-3β), inositol phosphate, and calcium-dependent cell survival signaling pathways. Lithium ions modulate serotonin receptor activity, increase acetylcholine receptor activity, and regulate corticotropin-releasing factor gene expression (which affects catecholamine metabolism). Lithium protects neurons in the cerebellum, cerebral cortex, and hippocampus from glutamate excitotoxicity, which is also important in preventing distress. Glycogen synthase kinases are the most important lithium-targeting proteins in humans. GSK-3β regulates glycogen synthesis, synaptic plasticity, neuronal apoptosis, and the body’s circadian rhythm. Upon activation of adrenal and cortisol signaling under stress, excess GSK-3β activity inhibits CREB, β-catenin, and other transcription factors that promote the survival of neurons; however, inhibition of GSK-3β (in particular, by lithium ions) promotes it.
This paper presents the results of the systematization of data on the relationship between the activity of GSK-3 kinases, lithium, mechanisms of chronic stress formation, and stress response. 335 publications were found in the PUBMED database using the queries “lithium AND (Psychological stress OR distress)” and “GSK-3 AND (Psychological stress OR distress) NOT lithium”. Furthermore, we reviewed the involvement of GSK-3 in the molecular mechanisms of stress, the role of lithium ions in the inhibition of stress effects, and the prospects for adaptogenic applications of lithium ascorbate, a low-toxic organic salt of lithium. GSK-3 β and molecular mechanisms of stress GSK-3 kinases (i.e., proteins that phosphorylate other proteins) phosphorylate more than 40 proteins, among which are β-catenin, axins (which modulate β-catenin interactions in the Wnt cascade), MAP1B, MAP2, and CREB signaling proteins, hypoxia response factor HIF1, τ-protein, insulin receptor substrate (IRS1), myelin basic protein, nerve growth factor (NGF), proinflammatory transcription factor receptor NF-κB (p65 and p105), and Notch signaling protein. By affecting the proliferation, differentiation, and survival of T cells, GSK-3 α- and β-kinases are also key regulators of the balance of pro- and anti-inflammatory cytokine biosynthesis in the central and peripheral nervous systems. In particular, signal transmission through the GSK-3β-NF-κB T-cell cascade is an integral component of the body’s response to chronic stress. In a model of acute electric shock-induced stress in mice, administration of a selective GSK-3β inhibitor (ARA01) or a selective NF-κB inhibitor (DDTC) reduced behavioral changes induced by acute stress. The specific GSK-3β inhibitor AR-A014418 exhibited anti-stress effects during immobilization stress in mice induced by short-term (30 min) or long-term (120 min) immobilization. NF-κB is a mediator of the GSK-3β signaling cascade, so decreased levels of the phosphorylated (i.e., inactivated) form of GSK-3β and increased levels of the phosphorylated (i.e., activated) NF-κB factor are associated with the formation of an acute behavioral response to stress. GSK-3β kinase itself, in turn, is an important signaling protein that is phosphorylated by Akt kinase (protein kinase B) such that GSK-3β activity is significantly reduced by Akt [16]. The participation of GSK-3β in the acute stress response is associated, in particular, with a change in the phosphorylation state of this enzyme. Accordingly, exposure to acute stress can suppress Akt activity, which, in turn, can decrease GSK-3β phosphorylation, thereby increasing its activity (Fig. 1). In the experiment, it was shown that, for example, immobilization stress suppresses Akt activity, activates GSK-3β, and impairs the induction of long-term potentiation in the hippocampus. GSK-3β is involved in the realization of the stressor effects of glucocorticoid stress hormones. For example, experimental prenatal stress significantly increases the concentration of the inactive form of GSK-3 (phospho-Ser9-GSK-3β) in the frontal cortex, so that GSK-3β is an important intracellular target of the stressor effects of glucocorticoids on frontal cortex neurons. GSK-3β hyperactivity is an important factor in the pathogenesis of depressive disorders.
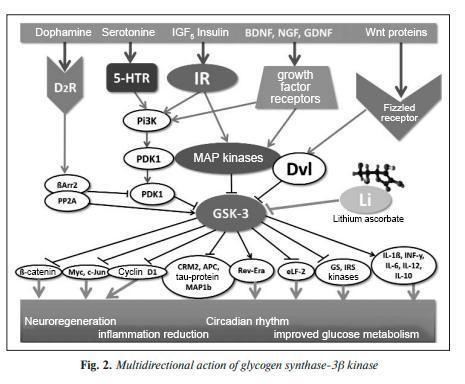
Overexpression of GSK-3β in the dentate gyrus of the hippocampus in mice caused effects similar to those of depressant substances and aggravated chronic stress. Administration of a selective serotonin reuptake inhibitor (fluoxetine) inhibited these effects and the apoptosis of hippocampal neurons. GSK-3β affects ion channels that realize the action potential of neurons. Mild but unpredictable chronic stress alters synaptic plasticity via GSK-3β-dependent modulation of Kv4.2 potassium channels. Recordings of channel activity on the surface of medium-spiking neurons showed that inactivation of the GSK-3β gene in the contiguous striatum nucleus reduced depression-like behavior and the severity of impaired long-term potentiation in the hippocampus. Interestingly, the level of GSK-3β gene expression in neurons directly correlates with behavioral abnormalities under stress. For example, individual differences in swimming test behavior in mice can predict the level of GSK-3β expression in neurons. The results point to the activation of GSK-3β expression in the brain when the contextual conditioning of unfavorable memories is increased, which is associated with a predisposition to depression. GSK-3 kinase β , neurotrophin BDNF, lithium, and GSK-3β stress counteraction are involved in the effects of many signaling molecules involved in the pathogenesis of distress: dopamine, serotonin, neurotrophic factors BDNF, NGF, GDNF, and others. (Figure 2). In particular, preclinical studies of rat brains have shown that inhibition of GSK-3β by lithium ions induces the expression of the BDNF neurotrophic brain factor. Note that one of the effects of clinical use of lithium is an increase in BDNF levels in the blood. BDNF promotes human neuronal growth through interactions with the Wnt/βcatenin signaling pathway, which is required for axon growth. GSK-3β, on the other hand, is an inhibitor of the Wnt cascade. When BDNF expression is elevated, the Wnt, Frizzled, Dsh, and β-catenin signaling factors are activated, and GSK-3β activity is reduced. On the contrary, when BDNF-dependent signaling was suppressed by microRNA (vs. BDNF gene mRNA), neuronal growth was reduced, components of the Wnt/β-catenin pathway were inhibited, and GSK-3β was activated. Inhibition of GSK-3β by lithium promotes activation of the Wnt neurotrophic cascade, accelerates differentiation of neuronal progenitor cells, stimulates astrocyte differentiation and myelin synthesis, supports neuronal survival, expression of neurotrophic factors, and others. The neurotrophic BDNF factor plays an important role in synaptic plasticity, neurogenesis, and neuronal survival by increasing the levels of proteins regulating mitochondrial biogenesis, protein quality control, neuronal resistance to oxidative, metabolic, proteotoxic, and alcoholic stress. BDNF is a neurotrophin that plays a significant role in cortical development, synaptic plasticity, neuronal survival and differentiation, and, consequently, in learning and memory. The BDNF neurotrophin BDNF is involved in the pathophysiology of chronic PTSD. Intracellular BDNF signaling is suppressed under conditions of hyperproduction of the stressor hormone glucocorticoids, an excess of which impairs synaptic plasticity, reducing spike density, neurogenesis, and long-term potentialization, effects associated with BDNF regulation by glucocorticoids. Excess GSK-3β activity is involved in the effects of chronic stress on nervous system development in rats. Prenatal stress has a broad impact on fetal outcomes, leading to increased expression of GSK-3β, which inhibits the expression of the supersonic hedgehog SHH protein involved in early embryogenesis, Wnt/β-catenin, and the brain neurotrophic factor BDNF [29]. Emotional distress in mice induced by exposure to variable frequency ultrasound is associated with the development of neuroinflammation and impaired hippocampal plasticity. Exposure to ultrasound altered the expression of genes related to serotonin metabolism and increased behavioral depression and/or aggression. A decrease in the density of dividing Ki67-positive and DCX-positive cells in the subgranular zone of the hippocampus and changes in the expression of the brain neurotrophic factor BDNF and its receptor TrkB were observed in mice exposed to stress. Also, mice exposed to the stressor showed increased levels of IL1β, IL6 in the hippocampus and blood plasma, increased activity of GSK-3β [30] and functionally related to GSK-3β proteins FOXO3a and PTEN against the background of a decreased phosphorylated (i.e., activated) form of the protein kinase Akt-phospho-Ser473 (Fig. 3). By inhibiting signaling in Erk-Creb-BDNF cascades, GSK-3β kinase activity exacerbates symptoms of depression in chronically stressed mice. Inhibition of GSK-3β leads to a reduction in depressive symptoms manifested in the open field test, tail suspension test, and forced swimming test by improving signaling from the BDNF receptor. In preclinical in vivo and in vitro studies, it was shown that lithium increases the expression of BDNF and other neurotrophins involved in neuronal survival and plasticity, including nerve growth factor, by inhibiting GSK-3β. Lithium is directly inhibited by lithium ions in the inhibition of stress effects The GSK-3β enzyme (GSK-3β gene), which is one of the main mechanisms by which the pharmacological effects of lithium salts are exerted. Lithium ions inhibit GSK-3β through competitive displacement of Mg2+ ion [32], and this effect is characteristic only for Li+ ions and is not observed for ions of other alkali metals (Na+, K+, Cs+, Rb+). Many physical and chemical properties of the Li+ ion are much closer to the properties of the Mg2+ ion (group IIA of the periodic system of elements of D.I. Mendeleev) than to the properties of ions.
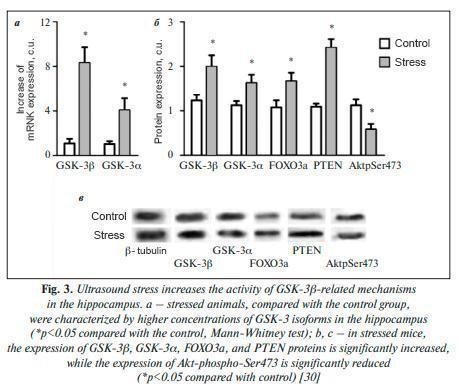
Alkali metals (IA group). In particular, Li+ and Mg2+ ions are non-polarizing “hard” cations with high charge density and strong affinity for oxygen-containing ligands.
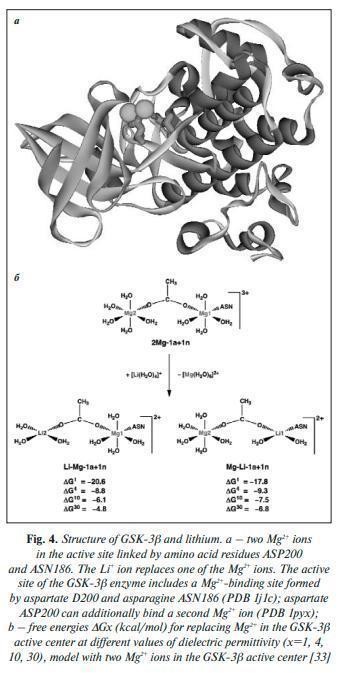
Both ions are characterized by close ionic radii: ri (Li+)=0.59 Å, and ri (Mg2+)=0.57 Å for coordination number of 4, ri (Li+)=0.76 Å and ri (Mg2+)=0.72 Å for coordination number of 6 [32]. In the work by T. Dudev and C. Lim [33], the coordination chemistry of Li+ and Mg2+ ions in the active center of GSK-3β was systematically analyzed by calculating the displacement energy of the Mg2+ ion by the Li+ ion. The results of the analysis showed that the competition between Mg2+ and Li+ ions depends on the total charge of the protein-cation complex, the number of metal cations, and the presence of a certain configuration of negatively charged groups in the active center. The unique configurations of GSK-3β and IMPA1 active centers guarantee that Li+ ion will inhibit these enzymes but not many other Mg2+-dependent enzymes (Fig. 4). The results of GSK-3β active center energetic simulations showed that when the GSK-3β active center binds two Mg2+ ions, replacing one of the Mg2+ ions with a Li+ ion becomes energetically advantageous. In addition to direct “structural” inhibition of GSK-3β, lithium ions can inhibit GSK-3β indirectly by increasing the phosphorylation of the amino acid Ser9 in GSK-3β via protein kinase B (PKB), which thereby inactivates GSK-3β [34]. Taking into account the previously described pathophysiological roles of GSK-3β in the formation of distress, it can be said that lithium directly affects the pathophysiology of stress. In particular, lithium-induced modulation of prefrontal noradrenergic metabolism stabilized the behavior of rats under chronic stress. Lithium affects noradrenergic signaling in the prefrontal cortex in chronically stressed rats. Lithium administration in chronically stressed rats reduced elevated levels of NET (membrane norepinephrine transporter) and VMAT2 (critical role in maintaining catecholamine levels in the central nervous system) proteins and MOA monoxidase enzyme activity to levels found in non-stressed rats. In addition, lithium reduced noradrenaline concentrations (by 24%) and shortened the period of immobility in animals subjected to immobilization stress. Lithium ions affect dopamine homeostasis, serotonin receptor activity, increase GABA receptor levels, attenuate NMDA receptor-activated signaling cascades, and inhibit cannabinoid dependence through modulation of cAMP, ERK1/2, and GSK-3β signaling pathways [36]. Lithium is able to block signaling from D2-dopamine receptors through inhibition of Akt/β-appectin-2/PP2A protein complex formation, which is necessary for GSK-3β activation [37]. In the experiment, the effectiveness of lithium salts application on different models of stress was shown. Lithium attenuated distress in mice induced by opioid receptor blockade (naltrexone) and assessed by forced swimming, tail suspension, and open-field tests. The effectiveness of lithium in chronic mild stress in rats exacerbated by neuroinflammation (injection of bacterial lipopolysaccharides) was demonstrated: lithium prevented the development of neuroinflammation and tauopathy (a neurodegenerative disease associated with abnormal τ-protein clumping into neurofibrillary tubules in brain tissue) and supported neuronal survival and preservation of cognitive function. In the C57Bl/6N mouse line with chronic psychosocial stress, lithium normalized the expression of proteasome proteins, oxidative phosphorylation, and antioxidant defense. Lithium (10 mg/kg) attenuated the proconvulsive and proinflammatory effects of 4-week social isolation stress in mice by engaging the nitrergic system. Co-administration of lithium (3 mg/kg) with nitric oxide synthase inhibitors inhibited the effect of stress on seizure amplification. Lithium blocked stress-induced depressive behavior (forced swimming) in rats by regulating the survival of hippocampal neurons through decreased GSK-3β activity. Stress led to increased corticosterone secretion and induced depression-like symptoms. The acute stress response was accompanied by a decrease in the division and differentiation of hippocampal cells and an increase in their apoptosis rate. Application of lithium salt or a specific GSK-3β inhibitor (AR-A014418) prevented the stress-induced increase in GSK-3β expression and decreased expression of synapsin-1 and BAG-1 protein in the hippocampus. Lithium also attenuated the stress-induced impairment of the long-term potentialization of hippocampus-dependent memory. An important component of the anti-stress action of lithium is its antioxidant and cytoprotective effects. Lithium supports the hippocampal antioxidant defense system in rats under conditions of chronic stress. Application of lithium salts in animals under stress conditions increased the activity of the enzymes glutathione peroxidase and glutathione reductase, catalase, heme oxygenase, decreased the concentration of malonic dialdehyde, and increased the expression of tyrosine hydroxylase. Lithium also decreased the activity of the monoamine oxidase B enzyme and increased hippocampal dopamine concentration to the level of unstressed animals. One of the key mechanisms of the antioxidant effect of lithium is stimulation of the expression of the Nrf2 transcription factor, which triggers the transcription of a number of genes containing “ARE” (Antioxidant Response Element) DNA sequences to prevent hyperactivation of oxidative processes. The cytoprotective effect of lithium salts has been found not only for nervous tissue but also for heart, kidney, and liver cells. Lithium salts are able to enhance autophagy in many neurodegenerative diseases by removing toxic protein aggregates, improving mitochondria, and also promoting hippocampal neurogenesis. Therapeutic concentrations of lithium significantly reduce levels of insoluble τ-protein and block the production of β-amyloid peptides pro Alzheimer’s disease by inhibiting GSK-3β. Lithium affects circadian rhythms by directly interfering with the function of the “gene clock” in the main rhythm-forming center of the brain, the suprachiasmatic nuclei of the hypothalamus. Lithium normalized circadian patterns of baseline emotional reactivity in mice under stress. These effects are most likely related to the inhibition of GSK-3β (which causes dysregulation of biorhythms), as well as to the possible effect of lithium on the production of the epiphyseal hormone melatonin (which is characterized by rhythm-stabilizing properties) [48, 49]. Lithium may enhance the effects of analgesics. For example, the addition of lithium to ketamine enhances insulin receptor signaling and the antidepressant effect of ketamine in a model of pharmacoresistant depression in rats. Animals were injected with adrenocorticotropic hormone (100 μg/day intraperitoneally, 14 days) and then treated for 2 days with ketamine (10 mg/kg), lithium (37 mg/kg), ketamine + lithium, or placebo (isotonic sodium chloride solution). According to the results of the “open field” and “forced swimming” tests, rats that received ketamine + lithium showed a more sustained antidepressant response (significant reduction in immobility time and latent immobility period).
Clinical and epidemiologic studies have shown that reduced levels of lithium in tap water are associated with psychotic experiences and distress in adolescents. Information on psychotic experiences, distress, and depressive symptoms was collected from adolescents attending public high schools in Japan (n=3040). Levels of lithium in tap water were inversely related to the incidence of psychotic experiences in adolescence (p=0.021). Lithium preparations were least characterized by the withdrawal syndrome, usually observed after discontinuation of psychotropic drugs. Among the drugs studied (benzodiazepines, non-benzodiazepine benzodiazepine receptor agonists, antidepressants, ketamine, and antipsychotics), selective serotonin reuptake inhibitors, serotonin and norepinephrine reuptake inhibitors, and antipsychotics were also consistently associated with persistent post-abstinent disorders. When lithium medications were discontinued, distress was practically absent.
The prospects of “anti-stressor” application of lithium ascorbate Lithium ascorbate is a pharmaceutical form of lithium that is very promising from the point of view of adaptogenic applications. Firstly, it is one of the least toxic lithium salts: for white rats of the Wistar line, the LD50 of lithium ascorbate was 6334 mg/kg, LD100 – 8000 mg/kg. Thus, according to the drug distribution table, lithium ascorbate belongs to the 5th class of “practically non-toxic” (LD50 ≥5000 mg/kg). Compared, for example, with the ubiquitous lithium carbonate (LD50 – 531 mg/kg), lithium ascorbate is 12 times less toxic. Secondly, chemo reactor modeling of lithium ascorbate effects in comparison with other organic lithium salts showed that the ascorbate anion, in comparison with control molecules (nicotinate, oxybutyrate, and comenate), has a higher affinity for dopamine, serotonin, benzodiazepine, and adrenergic receptors. Higher affinity to receptors indicates the possibility of modulation of the activity of these receptors by lithium ascorbate and more intensive transport of lithium ascorbate inside neurons. Thirdly, the ascorbate anion is characterized by independent action, complementing the effects of lithium ions. Ascorbate anion can exhibit anxiolytic, moderate anticoagulation, antihyperlipidemic, and antihyperglycemic effects. The protective effect of ascorbic acid on the behavior of stressed mice has been demonstrated. Acute immobilization stress induced depressive-like behavior in the forced swimming test. Oral administration of ascorbic acid (1 mg/kg) or fluoxetine (10 mg/kg) 1 h before stressor exposure prevented the stress-induced increase in immobility time in the forced swimming test. In addition, ascorbic acid reduced lipid peroxidation to control levels by restoring superoxide dismutase, glutathione reductase, and glutathione peroxidase activities in the cerebral cortex and in the hippocampus. Fourth, the in vitro study showed that in the concentration range of 0.1-1.0 mM lithium ascorbate exhibited a neuro-cytoprotective effect against the cytotoxic effect of glutamate. In in vivo experiments in rats, the drug contributed to the preservation of the pool of eosinophils, and reduction of adrenaline and noradrenaline levels in blood and improved the adaptation parameters of animals in the suspension test and in the model of transport stress. Lithium ascorbate was also effective for protection of myelin sheaths and differentiation of oligodendrocytes of the nervous system against alcoholic and oxidative (glutamate) stress. Conclusion. The adaptogenic and anti-stressor properties of lithium are manifested through several complementary mechanisms leading to an increase in neuronal viability/function, enhancement of neurogenesis, and maintenance of stress-induced homeostasis. The results of experimental and clinical studies of lithium salts indicate the prospect of their use to improve stress resistance. Lithium ions are natural inhibitors of excess activity of both GSK-3 isoforms, which promotes not only normothymic and antidepressant effects, but also anti-stressor effects of lithium. Lithium ascorbate supplementation is a safe and effective way to replenish the insufficient lithium supply that often accompanies distress.
It should be noted that the anti-stressor effects of lithium may be important for the prevention of alcohol dependence, whose mechanisms of formation include dysfunctions of dopaminergic and other mediator systems of the brain and humoral and neurotrophic factors on which lithium acts. The Normotim drug, which is based on lithium ascorbate, is also used in the complex treatment of alcoholism.




